
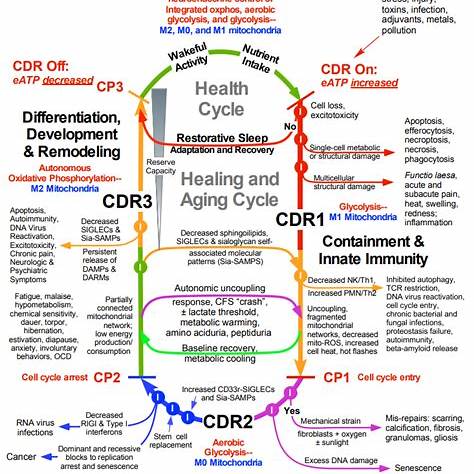
Resolving the Cell Danger Response:
A Multi-Pronged Protocol for Neuroimmune and Mitochondrial Recovery
Author: Noah El-Bermani
⚠️ Disclaimer
This post is a theoretical exploration of biochemical and immunological pathways associated with the Cell Danger Response (CDR). It is not clinically validated and is not intended as medical advice. No human or animal studies have been conducted using this exact protocol. Any implementation of these ideas should be done under the guidance of a qualified medical professional.
I. Removing or Neutralizing the Initial Insult
Environmental or microbial insults—such as endotoxins, heavy metals, and persistent infections—can initiate the CDR through pattern recognition receptors (e.g., TLRs). These insults disrupt redox balance and mitochondrial membrane potential, leading to inflammatory signaling and purinergic activation.
Interventions:
- Chelation (e.g., DMSA, EDTA): only for confirmed toxicity; otherwise consider chlorella as a food-based alternative2.
- Liver support:
- Binders: activated charcoal and bentonite clay to bind endotoxins.
- Autophagy enhancers: fasting, spermidine (1–2 mg), or berberine (200–400 mg)6.
II. Degrading Extracellular ATP (eATP)
eATP is a ‘danger signal’ that activates P2X7 and NLRP3 inflammasomes7. It induces calcium influx, potassium efflux, and the release of IL-1β and IL-18, sustaining inflammation.
Strategies:
- CD39/CD73 pathway support: promotes degradation of eATP to adenosine, an anti-inflammatory molecule8.
- Hormetic stressors: heat (e.g., sauna), cold exposure, and thermal cycling increase heat shock proteins (HSP70, HSP27) which stabilize membranes and reduce DAMP signaling9.
- Peptides:
- BPC-157 (250–500 mcg): supports endothelial and mitochondrial health10.
III. Calming the Inflammasome & Restoring Mitochondrial Function
Damaged mitochondria release ROS and mitochondrial DNA, fueling inflammasome activation.
Therapeutic Tools:
- NLRP3 inhibitors: e.g., MCC950 (experimental)11.
- Peptides:
- Metabolic Reprogramming:
IV. Restoring Signaling Without Blocking Receptors
Unlike suramin (a broad purinergic antagonist), these interventions aim to normalize rather than block signaling.
Strategies:
- CD73 enhancement: boosts conversion of AMP to adenosine.
- Low-dose naltrexone (LDN): modulates TLR4 and microglial activity17.
- Polyphenols: resveratrol and luteolin inhibit NF-κB and stabilize mast cells18.
Summary
This layered protocol targets multiple checkpoints in the CDR cascade. While unproven, it integrates concepts from immunology, redox biology, and mitochondrial medicine to theoretically restore cellular homeostasis. Any future experimental use should be preceded by rigorous clinical study.
References
- Naviaux, R. K. (2014). Metabolic features of the cell danger response. Mitochondrion, 16, 7–17. ↩
- Queiroz, M. L. S. et al. (2003). Protective effects of chlorella vulgaris in lead-exposed mice. Phytotherapy Research, 17(7), 675-680. ↩
- Dinkova-Kostova, A. T., & Talalay, P. (2008). NAD(P)H:quinone acceptor oxidoreductase 1 (NQO1) as a marker for cancer chemopreventive agents. The Journal of Biological Chemistry, 283(7), 3949-3955. ↩
- Pizzorno, J. (2014). Glutathione! Integrative Medicine: A Clinician’s Journal, 13(1), 8–12. ↩
- Lucock, M. (2000). Folic acid: nutritional biochemistry, molecular biology, and role in disease processes. Molecular Genetics and Metabolism, 71(1-2), 121-138. ↩
- Eisenberg, T. et al. (2016). Cardioprotection and lifespan extension by the natural polyamine spermidine. Nature Medicine, 22(12), 1428–1438. ↩
- Di Virgilio, F. et al. (2017). Extracellular ATP and P2 purinergic signalling in the tumour microenvironment. Nature Reviews Cancer, 18(10), 601–618. ↩
- Antonioli, L. et al. (2013). Anti-inflammatory effects of adenosine signaling in intestinal inflammation. British Journal of Pharmacology, 169(2), 302–312. ↩
- Chung, J. et al. (2008). Induction of HSP70 and HSP27 by sauna: role in cellular protection. Journal of Thermal Biology, 33(8), 422–428. ↩
- Sikiric, P. et al. (2018). BPC-157 and the central nervous system. Current Neuropharmacology, 16(6), 813–828. ↩
- Coll, R. C. et al. (2015). A small-molecule inhibitor of the NLRP3 inflammasome for the treatment of inflammatory diseases. Nature Medicine, 21(3), 248–255. ↩
- Lee, C. et al. (2015). The mitochondrial-derived peptide MOTS-c promotes metabolic homeostasis and reduces obesity and insulin resistance. Cell Metabolism, 21(3), 443–454. ↩
- Zhao, K. et al. (2004). A targeted antioxidant that protects mitochondria. The Journal of Biological Chemistry, 279(33), 34682–34690. ↩
- Hashimoto, Y. et al. (2001). Humanin inhibits neuronal cell death by interacting with a neuron-specific pro-apoptotic protein. Nature, 412(6848), 395–399. ↩
- Gomes, A. P. et al. (2013). Declining NAD+ induces a pseudohypoxic state disrupting nuclear-mitochondrial communication during aging. Cell, 155(7), 1624–1638. ↩
- Puchalska, P. & Crawford, P. A. (2017). Multi-dimensional roles of ketone bodies in fuel metabolism, signaling, and therapeutics. Cell Metabolism, 25(2), 262–284. ↩
- Younger, J. et al. (2014). Low-dose naltrexone for the treatment of fibromyalgia. Pain Medicine, 15(5), 867–875. ↩
- Weng, Z. et al. (2012). Luteolin suppresses mast cell-mediated allergic inflammation by inhibiting the MAPK and NF-κB pathways. Molecular Nutrition & Food Research, 56(9), 1414–1423. ↩
{kind=link}